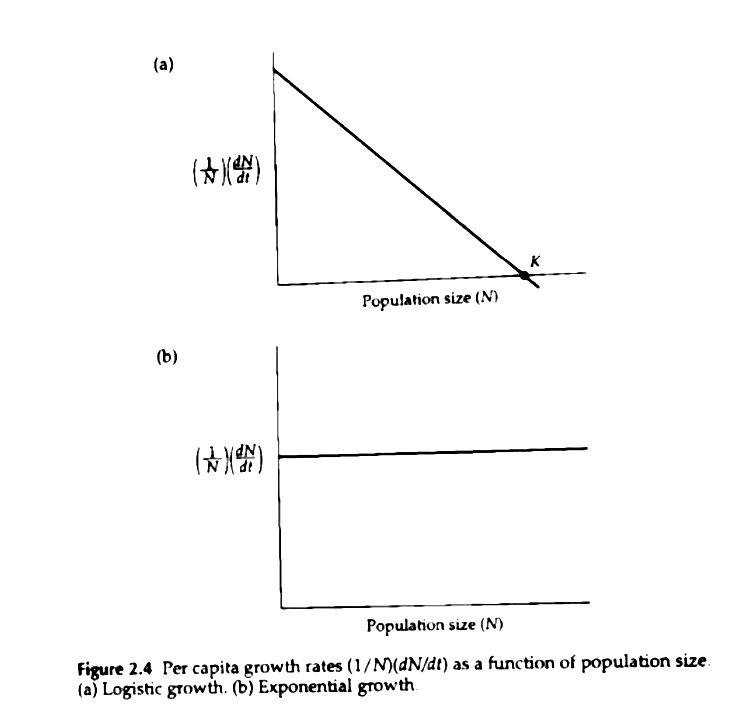
 The change in per capita population growth rates as a function of population size. from Gotelli "A Primer of Ecology". Density dependence is shown by the negative slope in the top graph. The decline in population growth rate as the population grows, gives the logistic population growth curve its distinctive S-shape. Density dependence, or a correlation of per capita population growth rate with population size, is a biological phenomenon that is ubiquitous in nature. Density dependence may either be positive or negative, depending if ∆N(b-d), increases or decreases as population size (N) increases (where b and d are births and deaths, respectively). Cases of positive density dependence (i.e., Alee effects) are rare in nature, however Negative Density Dependence (NDD; sometimes called a Malthusian-Darwinian process) is quite common. Since the first basic identification that the human population is limited by food production (Malthus 1798), the study of density dependence has become increasingly nuanced. The nuances of density dependence and it workings stem from the fact that it is impossible to identify process (i.e., the mechanism underlying density dependence) solely from pattern (i.e., a time series of population counts), and density dependence can be difficult to detect. Long time-series data at the appropriate scale for population fluctuations of the study organism are needed.
A classic example of how density dependence operates, can be understood from the data presented by Davidson and Andrewartha (1948) on Thrips imaginis and their host plants. Davidson and Andrewartha sought to describe the interannual variability in the population growth of the thrips using an exponential (i.e. density independent) growth function. Although, the fit of the exponential growth function described the data relatively well, Frederick Smith (1961) incorporated the inherit seasonality in the data with a piece-wise (i.e., multiple) regression with slopes that varied with winter degree days and amount of spring day. He demonstrated that the population growth data for the thrips were highly seasonal (i.e., density dependent), with population growth distributed normally around November/December. This can be seen visually from the data table represented in the original paper by Davidson & Andrewartha. The thrips go through boom and bust cycles that are regulated by seasonality. Aside from thrips, NDD appears many times in the body of ecological literature. In tropical forest ecology, the classic example is the famous “Janzen-Connell” hypothesis (now more of theory). Around the boom of when population biology was in the renaissance of density dependence (i.e., the 1970s), Dan Janzen and Joe Connell were both walking around tropical forests, independently. They observed that seedling densities were highest immediately below the tree crown. This phenomenon can be attributed to dispersal limitation of seeds originating from the parent tree. Trees have many dispersal mechanisms including wind dispersal (amenochory), animal dispersal (zoochory), and mechanical dispersal (autochory). However, gravity dispersal is certainly the most common mode of dispersal by trees in that all seeds will fall to the ground in the absence of a seed disperser, demonstrating why most reproductive seeds do not fall from the parent tree. Janzen (1970) and Connell (1978) thought about density dependence and its implications for the maintenance of diversity, and came to the theoretical conclusion that that seedling survival increases with seed dispersal distance. The mechanism by which increased mortality of seedlings closer to the parent tree occurs is understood to be pathogen and/or herbivore related (Comita et al. 2010). One can envision how common species are more frequently consumed by generalist herbivores, which target seedlings directly beneath the parent tree. Therefore, greater dispersal distances result in higher rates of seedling survival, and thereby maintain species diversity in highly-diverse tropical forests. Furthermore, NDD in tropical forests can be a mechanism for population regulation for many species. Recent work (Lebrija‐Trejos et al. 2016; Uriarte et al. 2018) has focused on how NDD is mediated via water availability in tropical seedlings, resulting in interacting effects of moisture and biotic interactions (likely belowground fungi or pathogens) on seedling distribution patterns and diversity. At Barro Colorado Island in Panama, Conspecific NDD was stronger in years with higher soil moisture. Also, trees with larger seed mass exhibited decreased levels of conspecific NDD, suggesting that seeds with greater seed mass have weaker NDD-regulation (i.e., a less negative slope of the density dependence as population size increases. Similarly, at Luquillo, Puerto Rico, the positive effects of increased solar radiation on seedling survival and growth were more pronounced, favoring drought tolerant species (e.g., lianas) and light-demanding pioneers and palm seedlings. In conclusion, NDD is a strong regulating force in population dynamics for many organisms and should be examined considering abiotic and biotic drivers of population change. ♠ Comita, L. S., H. C. Muller-Landau, S. Aguilar, and S. P. Hubbell. 2010. Asymmetric density dependence shapes species abundances in a tropical tree community. Science 329:330-332. Connell, J. H. 1978. Diversity in tropical rain forests and coral reefs. Science 199:1302-1310. Davidson, J., and H. Andrewartha. 1948. Annual trends in a natural population of Thrips imaginis (Thysanoptera). The Journal of Animal Ecology:193-199. Janzen, D. H. 1970. Herbivores and the number of tree species in tropical forests. American Naturalist:501-528. Lebrija‐Trejos, E., P. B. Reich, A. Hernández, and S. J. Wright. 2016. Species with greater seed mass are more tolerant of conspecific neighbours: a key driver of early survival and future abundances in a tropical forest. Ecology Letters 19:1071-1080 Malthus, R. T. 1798. An Essay on the Principle of Population. As it affects the future improvement of society, with remarks on the speculations of Mr. Godwin, M. Condorcet, and other writers. London. Smith, F. E. 1961. Density dependence in the Australian thrips. Ecology 42:403-407. Uriarte M., R. Muscarella, and J. K. Zimmerman. 2018. Environmental heterogeneity and biotic interactions mediate climate impacts on tropical forest regeneration. Global Change Biology 24: e692-e704.
1 Comment
Luke Lamb
2/10/2018 10:17:47 pm
At the community level, Hubbell (2006) empirically modeled the evolution of ecological equivalence across species in BCI. Dispersal- and recruitment-limitation more strongly accounted for tropical tree species richness than density dependent-limitation. It's interesting to see how drivers at different levels of organization change. Cool post Leave a Reply. |

AuthorJames "Aaron" Hogan is an ecologist interested in plant biodiversity, forests and global change. Archives
November 2021
Categories
|

|

|